Midori TUDA's research activities, collaborators,
awards, and more
Diversity and conservation of seed-feeding insects (JSPS Grant-in-Aid for Scientific Research (C), 2007-2009 to MT)
Large
seed size is beneficial for germination. On the other hand, the benefit of
large seed size in terms of escape from predators is not straightforward: Seed
predators attack larger/smaller seeds, depending on plant-herbivore
combinations: Larger seed attackers are often free from interspecies
competition, whereas smaller seed attackers tend to have competitors. This is
probably because mothers pay more costs to find and deposit their eggs on
larger seeds in specialist bruchines and the cost may
be paid off in the next generation. Additionally, seed predator larvae in small
seeds are more prone to parasitoid attack. This is probably equivalent to what
happens to gall makers. Interestingly, seed germination is promoted by
predation in few plant species.
In East European plant (seeds of Vicia, Lathyrus and Pisum)
and herbivore (Bruchus seed predators)
interactions...
For plant (legume)-herbivore (seed predator) relationship to be mutually
symbiotic, the number of herbivore species per plant species had to be one
(independent of the herbivore being specialist/generalist), whereas other
factors like the plant being annual/perennial, the size, hardness and number of
seeds, the body size of herbivores, and ancestral states were not important. In
the most symbiotic combination of species, the germination of uninfested seeds was impossible, the number of eggs laid
per seed was the highest, and the probability of sympatry with other related
legumes was the lowest. Thus, since this legume species requires a symbiotic
herbivore for its germination and forest rim with few competitors, it is
relevant to conserve such habitat for its existence.
A braconid wasp, Heterospilus prosopidis
 From top: a female and a male H.
prosopidis (Braconidae), and a female and a male Callosobruchus
chinensis (Bruchinae)
From top: a female and a male H.
prosopidis (Braconidae), and a female and a male Callosobruchus
chinensis (Bruchinae)
The
female wasp is in attempt to oviposit on the surface of larvae or pupae of the
azuki bean beetle C. chinensis inside beans (see below). Several genera
of the bean beetles (Chrysomelidae: Bruchinae) are known as stored product
pests (e.g., Callosobruchus, Acanthoscelides, Caryedon
and Bruchidius) (Tuda, 2007).
Evolution of stored bean pests has been driven primarily by a climatic factor
(long dry season) (Tuda et al., 2006).
Conversely, some bean beetles are beneficial and used as control agents of
weedy plants (Tuda, 2007).
Ecological studies using bruchine beetles in the lab have been contributed to
the development of population and evolutionary dynamics theories.
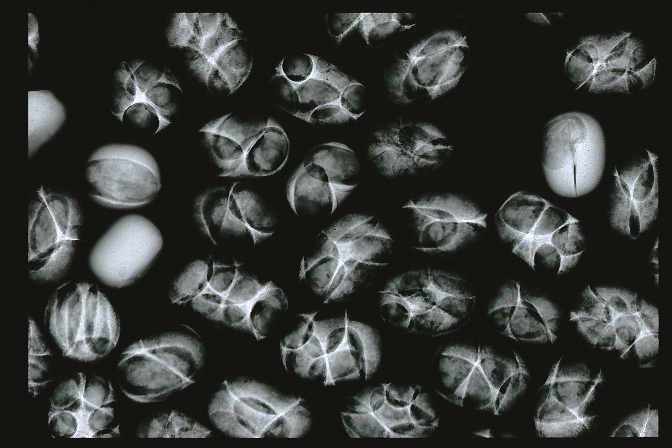
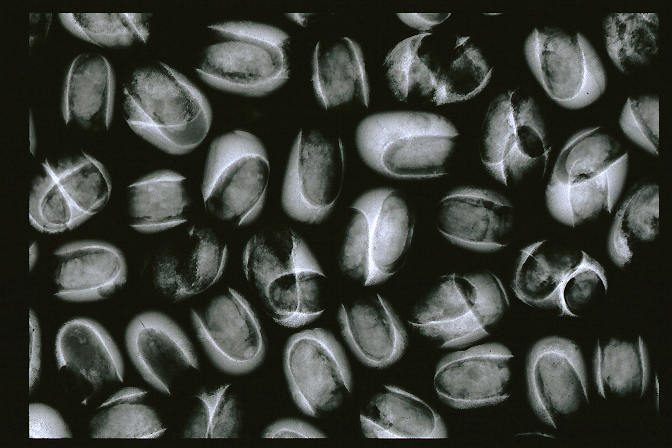
Bruchine larvae and pupae inhabiting inside beans (soft-X-ray images of infested beans)
Cavities made by bruchine larvae.
You can see the larvae and pupae inside the cavities.
|
Scramble-type larvae avoid competition |
Contest-type larvae kill each other |
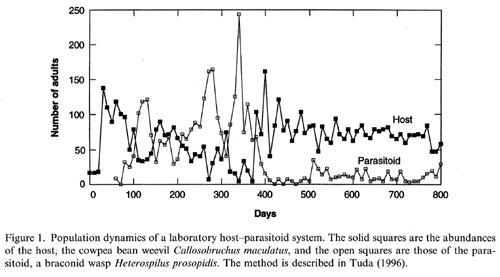
Evolutionary population dynamics of the cowpea bean beetle (the host) and Heterospilus prosopidis (the parasitoid)
The
larval competition type of the cowpea bean weevil has shifted from the scramble
type to the contest type after about 400 days, that is, 20 host generations.
The ecological characteristics of the parasitoid have not changed. We found
that the evolutionary change in the host competition type alone can generate
the stabilization and the switch in population levels between host and
parasitoid (Tuda and Iwasa,
1998; Tuda, 1998; Tuda and
Shimada, 2005).
Our evolutionary model shows that it was the bean size that increased the
frequency of the contest type.
This is one of the few studies that demonstrate an evolutionary change influences
population dynamics of both host and parasitoid.
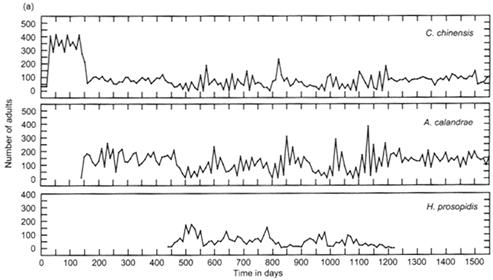
Introduction of an alien parasitoid and its ecological consequences
The
introduction of an alien parasitoid (on day 440) modified an ecological trait
of a native parasitoid, which consequently altered the dynamical behavior of
the whole ecological assemblage from stability to unstable complex dynamics (Tuda and Shimada, 2005).
From top; the host (Callosobruchus chinensis), the native parasitoid (Anisopteromalus
calandrae) and the alien parasitoid (Heterospilus prosopidis)
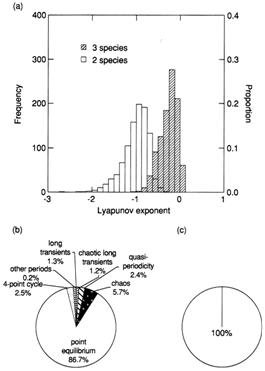
Before the introduction of the alien parasitoid (blank bars, right pied chart).
After the introduction (gray bars, left pied chart). Top: dominant Lyapunov
exponents of bootstrapped population dynamics. Some time series were chaotic
(indicated by positive Lyapunov exponents). Bottom: population behavior of the
bootstrapped time series.
Midori TUDA's list of papers, research activities,
collaborators, awards, and more
Link
to the e-bean
------------------------------------
Out of town:
4-12 July, 2012 First Joint Congress
on Evolutionary Biology (Canada)
17-27 Nov, 2011 Field work (Thailand)
19-22 Aug., 2011 International workshop on global legume
diversity assessments (Fukuoka)
12-28 July, 2011 Field work
and experiment (Hungary)
17-27 June, 2011 Second
Entomophagous Insects Conference (France)
8-12 March, 2011 Annual meeting of Ecological Society of Japan
(Sapporo)
18-21 Feb, 2011 Seminar (Shizuoka)
12-16 Feb, 2011 Field work (Hawaii)
11-21 Nov, 2010 Field work (Thailand)
15-20 Mar, 2010 Annual meeting of Ecological
Society of Japan (Tokyo)
2-6 Aug, 2009 Journal of Experimental
Biology Symposium: Survival in a Changing World (Awaji)
18-19 Oct, 2008 Annual Meeting of the
Society of Population Ecology (Tokyo)
17-23 May, 2008 Field work (Hungary)
14-17 March, 2008 Annual
meeting of Ecological Society of Japan (Fukuoka)
11-20 Feb., 2008 Experiment and field work (Thailand)
19-22 Oct, 2007 Symposium of
Society of Population Ecology (Sapporo)
18-19 Sep., 2007 Invited talk at International Workshop on
Ecological Informatics of Chaos and Complex Systems- Spectral Imaging for
Ecosystem Modelling (Tokyo)
26 June-27 July, 2007 Research
visit (Hungary)
26 May-2 June, 2007 Field work
(Hungary)