研究内容Research Topics
主要研究項目
- イネ種子貯蔵タンパク質の生合成・集積を制御する遺伝的機構
- ナショナルバイオリソースプロジェクト(NBRP)におけるイネ突然変異系統の整備
- 在来イネ遺伝子資源の保存と特性評価に関する研究
本分野では、作物のもつ有用な遺伝子資源の収集と作出および特性開発と保存・管理に関する遺伝・育種学的研究を主要課題としている。品種改良の素材には有用な遺伝子資源が必要で、幅広い遺伝変異をもつ世界各地の栽培種を広く収集するとともに、人為的に突然変異を誘発し、有用な遺伝子資源を作出し、その特性を明らかにすることが重要である。こうした目的のために、本分野では、つぎのような研究を進めている
1.イネ種子貯蔵タンパク質の生合成・集積を制御する遺伝的機構
植物種子は主に1種類の貯蔵タンパク質を蓄積するが、イネ種子は3種類の性質の異なる貯蔵タンパク質を蓄積する。それらの一つはグルテリンと呼ばれ、全タンパク質の60%以上を占め、11Sグロブリンとアミノ酸配列において相同性が高い難溶性のタンパク質である。さらにイネ種子はアルコール可溶性の貯蔵タンパク質であるプロラミンを約20%、塩可溶性のαグロブリンを約10%含んでいる。これらの貯蔵タンパク質は細胞内膜輸送系(エンドメンブレンシステム)によってプロテインボデイ(PB)に蓄積される。
すなわち、プロラミンは小胞体由来のプロテインボディI (PBI)に、グルテリンとグロブリンは液胞型プロテインボディII(PBII)に集積している(図1-1)。イネ貯蔵タンパク質の2種類のPBの輸送蓄積に関わる遺伝学的制御機構について、これまでに得た知見を紹介する。
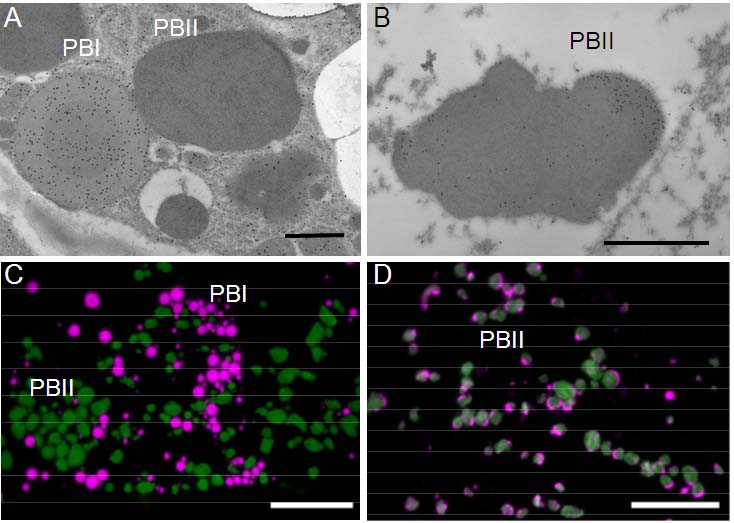
図 1-1. イネ胚乳におけるプロテインボディの免疫顕微鏡写真
A: プロラミンのPBIへの局在。B:
グルテリン(小さい金コロイド)とグロブリン(大きい金コロイド)のPBIIへの局在。C:マゼンタと緑はそれぞれプロラミンとグルテリン抗体による標識。D: 緑とマゼンタはそれぞれグルテリンとグロブリンによる標識。グロブリンはPBII内に偏在する。
1-1.プロラミンに関する突然変異
メチルニトロソウレアの受精卵処理によってプロラミンの蓄積に関するesp1、esp3、Esp4突然変異が作出された。esp1変異体では13 kDa
Cys-Pプロラミンの減少が、esp3変異体とEsp4変異体では、それぞれCys-Rプロラミンの減少と増加が認められた。野生型のPBIは中心部分の電子密度が大きく、PBの周囲に近づくに従って電子密度が小さくなり、同心円状構造を形成している。中心部にはCys-R分子種である10kDaプロラミンの集積により核を形成し、その周囲をCys-P分子種が取り囲んでいる
(図1-2)。esp3変異体では同心円状構造が見られず、内部の電子密度が均一であるプロラミンPBが観察された。Esp4変異体では、電子密度の大きい核を構成する部分の領域が広がり、中心部分が電子密度の大きなPBが観察された。野生型のPBIで認められる年輪上構造が認められなかったが、中心部の電子密度が高かった。esp1変異体では野生型と同じ年輪上構造が認められた。これらの結果は、10
kDa Cys-RプロラミンがPBIにおける核形成に重要であることを示唆している。また、esp1変異体は複数のプロラミン分子を減少させる変異であることを明らかにした(図1-3)。(Nagamine et al, 2011, Ushijima et al, 2011)
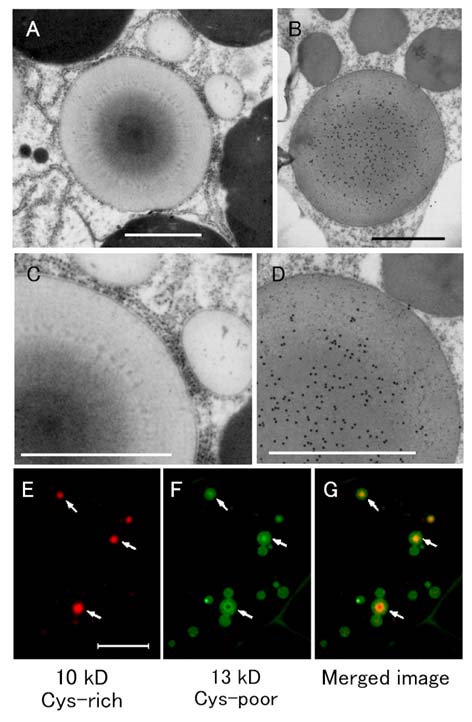
図 1-2. イネ胚乳におけるプロテインボディタイプI(PBI)の免疫電子顕微鏡及び免疫蛍光顕微鏡写真
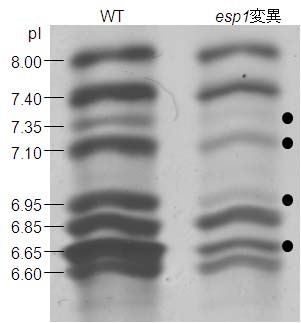
図 1-3. イネ種子プロラミンのIEF電気泳動像
esp1変異ではCysPプロラミンの内、●で示すバンドが減少した。
1-2. グルテリン前駆体を集積する突然変異
グルテリンの輸送から集積までの過程における遺伝的制御機構を明らかにするために、メチルニトロソウレアの受精卵処理によって、8系統のグルテリン前駆体を集積する突然変異esp2、Glup1、glup2、glup3、glup4、Glup5、glup6及びglup7を同定した。グルテリン、グロブリン及びプロラミンのタンパク質組成およびプロテインボディの形態に基づき、各変異をグルテリンの輸送過程に位置付けた(図1-4)。
その結果、小胞体内分別輸送に関与する遺伝子の変異としてesp2変異(クラス1変異)、小胞体からの輸送小胞形成に関与する変異としてGlup1、glup2及びglup7変異(クラス2変異)、小胞体から液胞までの輸送に関与する遺伝子の変異としてglup4、Glup5及びglup6変異(クラス3変異)、液胞内プロセシングに関与する遺伝子の変異としてglup3変異(クラス4変異)に分類した。
これまでの結果からクラス1に属する変異が最も上位に位置し、クラス2に属する変異はクラス3とクラス4に属する変異に対して上位であること、クラス3に属する変異はクラス4の変異に対して上位であること、各クラス内の変異は互いに相加的に作用することが明らかとなった(Ueda et al., 2010)。
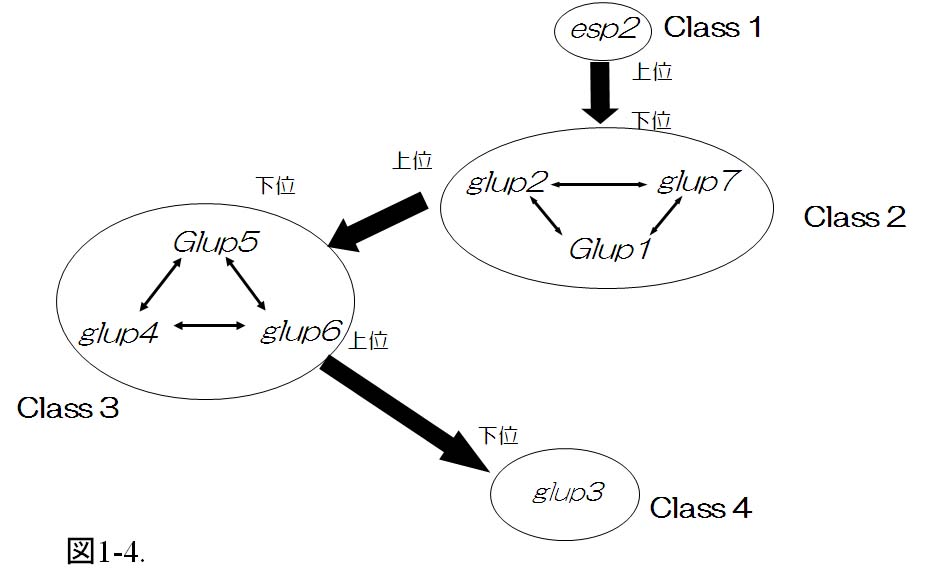
図 1-4. グルテリン前駆体を多量に集積する変異の遺伝子間相互作用
esp1変異がすべての変異に対して上位であった。Glup1、glup2、glup7変異がesp2変異の下位に位置しお互いに相加的であった。その下位にglup4、Glup5、glup6変異が位置していた。glup3変異がすべての変異に対して下位に位置した。
<小胞体内における修飾>
クラス1に属するesp2変異体において、タンパク質ジスルフィドイソメラーゼ(PDI)が欠損していた。esp2変異体における種子胚乳の電子顕微鏡観察によって、野生型と同等のPBIIが観察された。しかし、PBIは認められず、代りに直径約0.5μmの小胞体由来と考えられる顆粒が多数認められた(図1-5)。免疫組織化学的解析および生化学的解析によって、これらの顆粒中にはグルテリン前駆体とプロラミンが含まれており、両者は分子間ジスルフィド結合を形成していることが明らかとなった。小胞体内腔にグルテリン前駆体がプロラミンと結合した状態で集積していたことは、PDIは小胞体内における両タンパク質の分別に関与していること、及びPDIグルテリン前躯体の分子内ジスルフィド結合形成に関与していることを示している。しかしながら、グルテリン前駆体のPDIによる分子内ジスルフィド結合の形成が、当該タンパク質のPBIIへの輸送に必ずしも必要ではないと考えられる。すなわち、野生型登熟種子においてグルテリンのPBIIへの蓄積は開花後5日目ころから観察されるが、その時期はプロラミンの蓄積は認められない。esp2変異体の種子登熟初期において、グルテリン前駆体分子は正常に液胞に輸送されるが、開花後10日目頃において、新生プロラミンとグルテリン前駆体の凝集体が蓄積を開始する。これは、PDIの欠損によって、グルテリン前駆体とプロラミン分子内に適切なジスルフィド結合が形成されないため、非特異的分子間S-S結合が形成され、グルテリン前駆体とプロラミン凝集体を小胞体内に形成すると考えられる。(Takemoto et al 2002, Satoh-Cruz et al 2010)
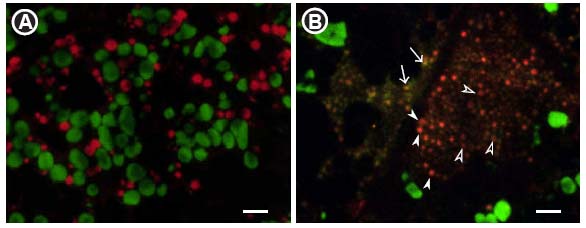
図 1-5. グルテリン前駆体を多量に集積するesp2変異の免疫蛍光顕微鏡写真
Protein
disulfide isomerase を欠損するesp2変異ではPBIIは観察されず、小胞体内にグルテリンとプロラミンが共存していた(B: 矢印、矢頭)。Aは野生型。赤と緑はそれぞれプロラミンとグルテリン抗体による標識を示す。
<ゴルジ体から貯蔵型液胞への輸送>
クラス3に属するglup4変異体においてグルテリン前駆体のPBIIへの輸送が阻害されていた。野生型で見られた直径2〜3μmのPBIIは認められず、それより小型でグルテリンを集積している貯蔵型液胞が観察された。さらに、野生型では認められない新奇構造体が観察され、そこにグルテリン前駆体が集積していた(図1-6)。この構造体には、小胞体タンパク質BipやPDI、ゴルジ体タンパク質糖鎖修飾酵素、細胞壁抗生物質βグルカン等の存在が認められた。この構造体には、小胞体タンパク質BipやPDI、ゴルジ体タンパク質糖鎖修飾酵素、細胞壁抗生物質βグルカン等の存在が認められた。同構造体は細胞膜と細胞壁の間のparamural
spaceに存在することからparamural body と命名した。連鎖解析によってGLUP4遺伝子は、GTPase活性を有し小胞輸送に関与する
Rab5aをコードしていることが明らかとなった。glup4変異体におけるグルテリン前駆体の構造体への集積は、Rab5aの機能の欠失によると考えられる。これらの結果からRab5aはグルテリン前駆体のゴルジ体から貯蔵型液胞への輸送に関与すると考察した。(Fukuda et al. 2011)。
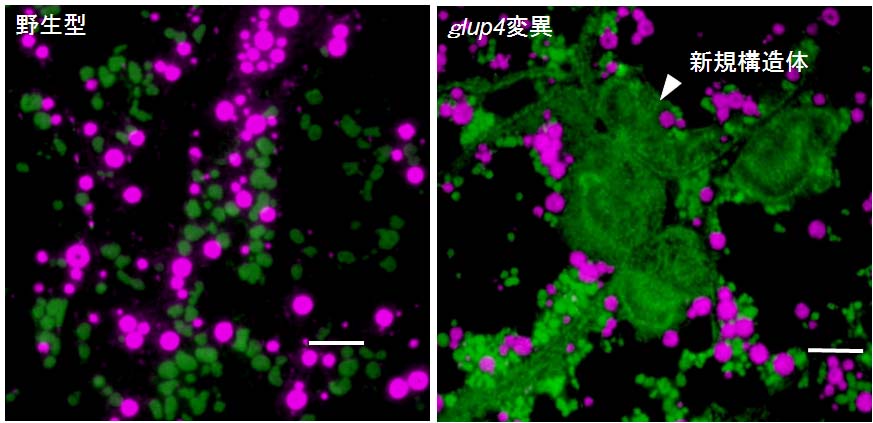
図 1-6. グルテリン前駆体を多量に集積するglup4変異の免疫蛍光顕微鏡写真
赤と緑はそれぞれプロラミンとグルテリン抗体による標識。Small
GTPase Rab5aを欠損するglup4変異では、野生型で見られない新規構造体内が観察され、同構造体にグルテリンが集積していた。
同じクラス3に属するglup6変異体の細胞内微細構造はglup4変異体と酷似していた。すなわち、glup6変異体とglup4変異体のPMBはほぼ同じ構造であり、グルテリン前駆体やゴルジ体タンパク質糖鎖修飾酵素、細胞壁抗生物質βグルカンを含んでいた。連鎖解析からGLUP6遺伝子はRab5タンパク質の活性化ドメインVPS9を有するグアニンヌクレオチド交換因子(guanine
nucleotide exchange factor: GEF)をコードしていることが明らかとなった。発現タンパク質を用いたin vitro
GEFアッセイの解析結果、GLUP6/GEFはGLUP4/Rab5の活性化因子であることが明らかとなった。これらの結果から、glup6変異体ではGLUP6/GEFの欠失に伴うRab5タンパク質の活性低下によりグルテリン前駆体のゴルジ体から貯蔵型液胞への輸送に支障を生じたと考察した。(図1-7)(Fukuda et al. 2013)
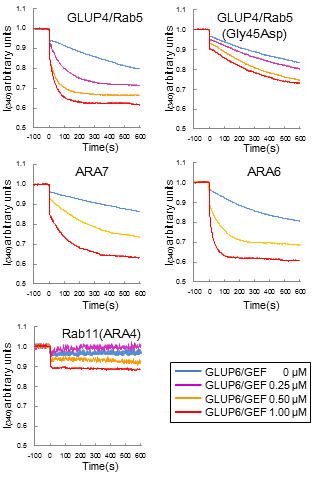
図 1-7. GLUP6/GEFによるGLUP4/Rab5のin vitro GEF assay GST- GLUP4/Rab5はGST- GLUP6/GEFによって活性化した。さらにシロイヌナズナのRab5ファミリーであるGST-ARA6とGST-ARA7も活性化した。これに対してシロイヌナズナのGST-Rab11タンパク質は活性化しなかった。これらの結果よりGLUP6/GEFはRab5ファミリーに特異的な活性化因子であることが明らかとなった。なお、GST- GLUP4/Rab5(Gly45Asp)タンパク質の活性はほとんど認められなかった。
<貯蔵型液胞内のプロセッシング>
クラス4に属するglup3変異では、グルテリン前駆体を貯蔵型液胞に集積していることが明らかとなった。この変異では貯蔵型液胞におけるグルテリン前駆体の切断が不完全であることを示している。同座のglup3変異3系統の登熟種子において、液胞タンパク質プロセッシング酵素(VPE)活性が野生型と比較して顕著に減少していた。これら3系統におけるVPE遺伝子では、野生型と比較してアミノ酸置換あるいは終止コドンの形成が認められた。さらに同変異体のPBIIでは、野生型のPBIIで観察されるグルテリンの結晶格子構造が認められなかった(図1-8)。これらの結果は、GLUP3遺伝子はVPEをコードしていること、VPEは貯蔵型液胞におけるグルテリン前駆体のプロセッシングに関与していること、VPEによるグルテリン前駆体のプロセッシングはグルテリンの集積の際に結晶構造形成に必須であることを示している。(Kumamaru et al, 2010)

図 1-8. グルテリン前駆体を多量に集積するglup3変異のPBIIの免疫電子顕微鏡写真
直径15nmと5nmの金コロイドは、それぞれグロブリンとグルテリンによる抗体によって標識されたことを示す。野生型ではグロブリンがPBIIに偏在している。グロブリンが偏在しない部位ではグルテリンの結晶格子構造が観察された。液胞プロセッシング酵素を欠損するglup3変異では、PBII内にグロブリンは均一に集積し、グルテリンの結晶格子構造は観察されなかった。
2. ナショナルバイオリソースプロジェクト(NBRP)におけるイネ突然変異系統の整備
イネの穂の変異体 |
ゲノム配列解読後のライフサイエンスの重要な研究課題は、遺伝子機能の解明である。遺伝子の機能を研究する方法の一つに、突然変異体を利用した解析がある。多くの生物種で塩基置換突然変異を高頻度で生じさせるN-methyl-N-nitrosourea (MNU)を突然変異原とする受精卵処理法を用いて、水稲品種「金南風」および「台中65号」を原品種とする形態突然変異系統をそれぞれ約3,000系統、合計約6,000系統整備した。 |
TILLING法を用いた解析の結果は、これらの変異系統プールはイネゲノム中のすべての遺伝子の突然変異を網羅していることを示唆している。イネバイオリソースのさらなる高品質化を図るため、利用者にとって利便性の高い、生育期間が短くグロースチャンバー内でも栽培・評価が可能なイネ突然変異プールの確立を目指して、北海道の水稲品種「キタアケ」や「ゆきひかり」を原品種とする形態突然変異系統を整備中である。
3. 在来イネ遺伝子資源の保存と特性評価に関する研究
|
|
新品種開発には,遺伝資源とその情報の収集・整理・管理が必要である。遺伝子資源であるイネの品種を保存・収集し、特性を評価することは、品種改良のための育種素材の提供に加え、イネの進化や、遺伝育種学、生理・生化学、分子生物学などの研究に重要な材料を提供する。 |
本研究分野では、温帯地域の各地から収集してきた約4,000点のイネ品種を保存するとともに、特性の評価を続けている。保存品種の中には、最早農家から得ることのできない貴重な遺伝子資源も含まれている。保存するイネ系統はデータベースとして公開している。(http://www.shigen.nig.ac.jp/rice/rice-kyushu/htdocs/main.html)