なぜ黄麹菌を研究に用いるか?
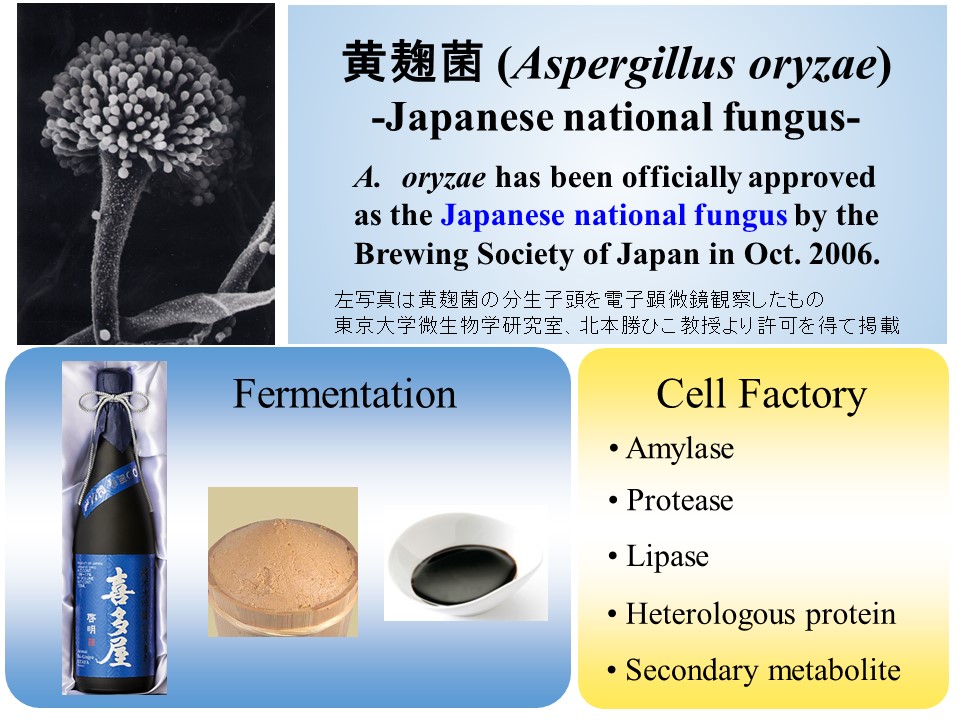
黄麹菌Aspergillus oryzaeは、古くから我が国において日本酒、味噌、醤油といった発酵・醸造産業に利用されてきた有用な微生物であり、安全性の高い菌としてGRAS(Generally Regarded As Safe)に認められています。そうした経緯から2006年10月、日本醸造学会大会において黄麹菌は、白麹菌、黒麹菌とともに国菌に認定されています。黄麹菌は、非常に高い有用物質分泌能力を有し、アミラーゼなどの有用酵素やコウジ酸といった有用二次代謝産物を菌体外に大量に分泌します。こうした性質を利用して、黄麹菌を用いて我々ヒトにとっての有用物質を安全かつ大量に生産させる研究が数多くなされています。さらに2019年10月にはゲノム編集食品が解禁されたことから、黄麹菌自体のことを細胞生物学的により一層理解し、ゲノム編集により黄麹菌を分子育種することで、発酵・醸造産業においても新たな展開が期待されます。
研究対象としての黄麹菌
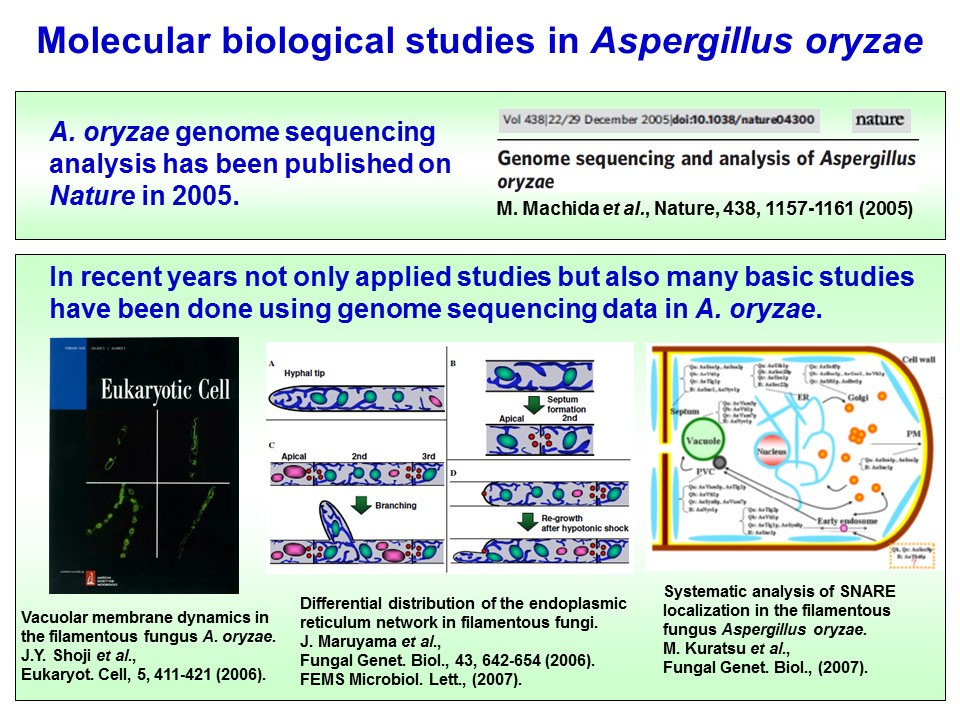
上述のように、黄麹菌は古くから有用菌種として知られていましたが、有性世代が確認されておらず、古典的な遺伝学的研究手法を用いることができない、多核の多細胞生物であるなどといった研究材料としては扱いにくい性質を有するため、長い間その研究は応用研究にとどまり、本格的な基礎研究はなされていませんでした。しかし、2005年に麹菌ゲノム解析コンソーシアムにより黄麹菌のゲノム解読が完了したことにより、その情報を利用した研究が容易に行えるようになりました(Machida et al., 2005)。それにより、分子生物学的手法が確立され、同様にゲノム解読がなされたNeurospora crassaやAspergillus nidulansと同様に、基礎研究の対象としてモデル糸状菌となっています。実際に、細胞生物学的解析手法により、黄麹菌ではこれまでにGFP(Green Fluorescence Protein; 緑色蛍光タンパク質)を用いた細胞内オルガネラの解析(Maruyama et al., 2006; Shoji et al., 2006)や、オルガネラマーカータンパク質の確立がなされています(Kuratsu et al., 2007)。
黄麹菌を用いた有用物質生産
では実際に黄麹菌を用いてさまざまな有用物質を容易に大量生産することができるのでしょうか?残念ながら話はそう単純ではありません。というのも一般的に、有用物質をコードする異種の遺伝子を黄麹菌に単に導入しただけでは、その生産量は黄麹菌本来の生産能力に比べて圧倒的に低いのです。その理由には色々と考えられますが、異種タンパク質生産の場合、翻訳や分泌の効率が悪い、また分泌された後に培養上清中でプロテアーゼによって分解されてしまう、といったことが挙げられます。そこで、そうした原因を改善することによって、より多くの有用タンパク質を生産できるようになったという研究も多くなされています(Nemoto et al., 2009; Yoon et al., 2010; Ohno et al., 2011; Yoon et al., 2011)。
どのように黄麹菌を用いてより有用物質を生産させるか? ~細胞生物学的解析によるアプローチ~
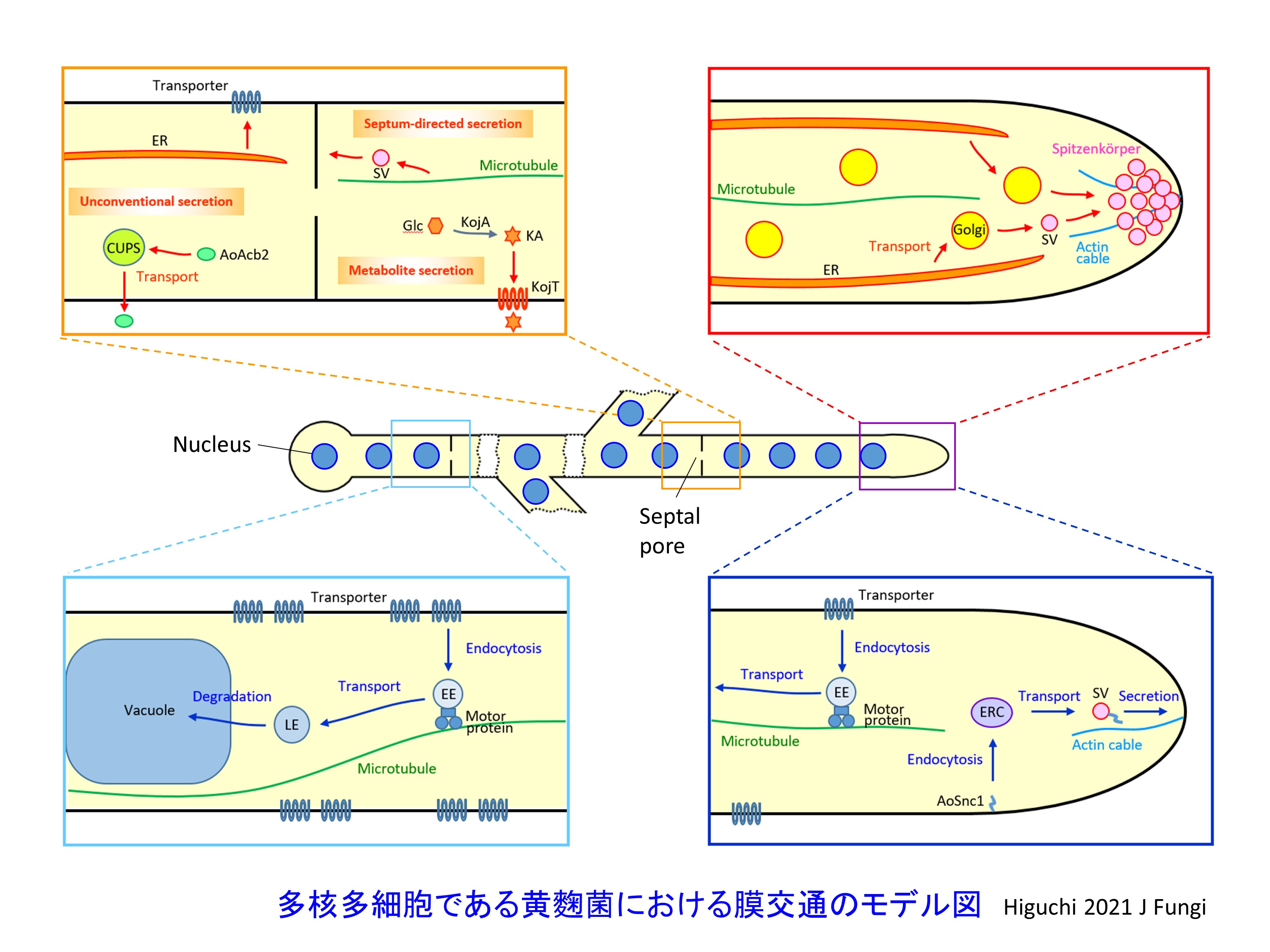
それでは今後、どのような戦略で有用物質を黄麹菌でより多く作らせることができるのでしょうか?我々はまず、細胞内における小胞輸送経路に着目します。分泌タンパク質は粗面小胞体のリボソームにより合成された後、ゴルジ体を経て細胞膜から細胞外に分泌されます。この際、タンパク質は脂質二重膜で覆われた小胞によって輸送されます。さらに、細胞内小胞輸送経路には分泌経路のみではなく、細胞膜から細胞内へと物質を取り込むエンドサイトーシス経路も含まれます。そして、細胞内小胞輸送経路は小胞による物質輸送だけではなく、脂質二重膜それ自体の細胞内でのバランスを調節するという意味でも重要です。そこで、細胞内小胞輸送経路全体をより詳細に解析し明らかにすることで、有用物質生産に対する新たなアプローチを見出そうとしています。
黄麹菌における膜交通の研究
膜交通(メンブレントラフィック)とは、小胞輸送経路を含めたその名の通り脂質膜を介したオルガネラ間における物質輸送経路のことを言います(Higuchi, 2021a,b)。黄麹菌を含む糸状菌において特徴的な膜交通の一例として、初期エンドソーム動態が挙げられます。エンドサイトーシス経路のオルガネラである初期エンドソームは、細長い菌糸と呼ばれる細胞の中を、毎秒約2マイクロメートルの速さで動態を示します。そしてこの初期エンドソーム動態が、タンパク質翻訳構造体であるポリソーム(複数のリボソームが結合したmRNA)の細胞内分布に重要であるということが明らかになりました(Higuchi et al., 2014; summarised in Higuchi and Steinberg, 2015)。さらに黄麹菌においては、初期エンドソーム動態がα-アミラーゼ生産や細胞分化に重要であることもわかってきました(Togo et al., 2017)。また一方で、タンパク質特殊分泌経路(Kwon et al., 2017)、分泌糖タンパク質改変(Li et al., 2020)、タンパク質品質管理機構(Morita et al., 2020)といった解析も行っています。
多核多細胞である黄麹菌における時空間発現制御機構
我々は近年、多核多細胞である黄麹菌において、α-アミラーゼやアクチン細胞骨格、EGFP融合タンパク質をコードするmRNAがどのような細胞内局在を示すのかを明らかにしました(Higuchi and Takegawa, 2020; Morita et al., 2021)。また、生きた細胞でmRNAを可視化できるMS2システムを適用することで、糖化に重要な酵素であるグルコアミラーゼをコードするglaA mRNA、微小管細胞骨格の構成タンパク質であるβ-チューブリンをコードするbtuA mRNA、小胞体ストレス依存的に発現誘導され小胞体シャペロンをコードするbipA mRNAの時空間的な発現と局在の分子制御機構を明らかにしました(Morita et al., 2024; Kawatomi et al., 2024; Sathongdejwisit et al., 2025)。
今後は、有用物質やその生合成に関連する因子をコードする遺伝子が、黄麹菌多核多細胞のどこでいつ発現し、タンパク質として作られているのかを解析することで、有用物質の生合成機構の全貌を時空間的に明らかにしていきたいと考えています。こうした膜交通経路におけるさまざまな分子機構を明らかにし、細胞内物質輸送の仕組みをうまく調節することで、有用物質をより多くまたもしくはバランス良く生産する黄麹菌の育種を目指しています。