
植物生産生理学研究室では主に以下の研究を行っています。
英語版はこちら。The English version is here.
光合成型・ストレス耐性機構の改変による環境ストレスレジリエント(強靭)な新規作物の作出
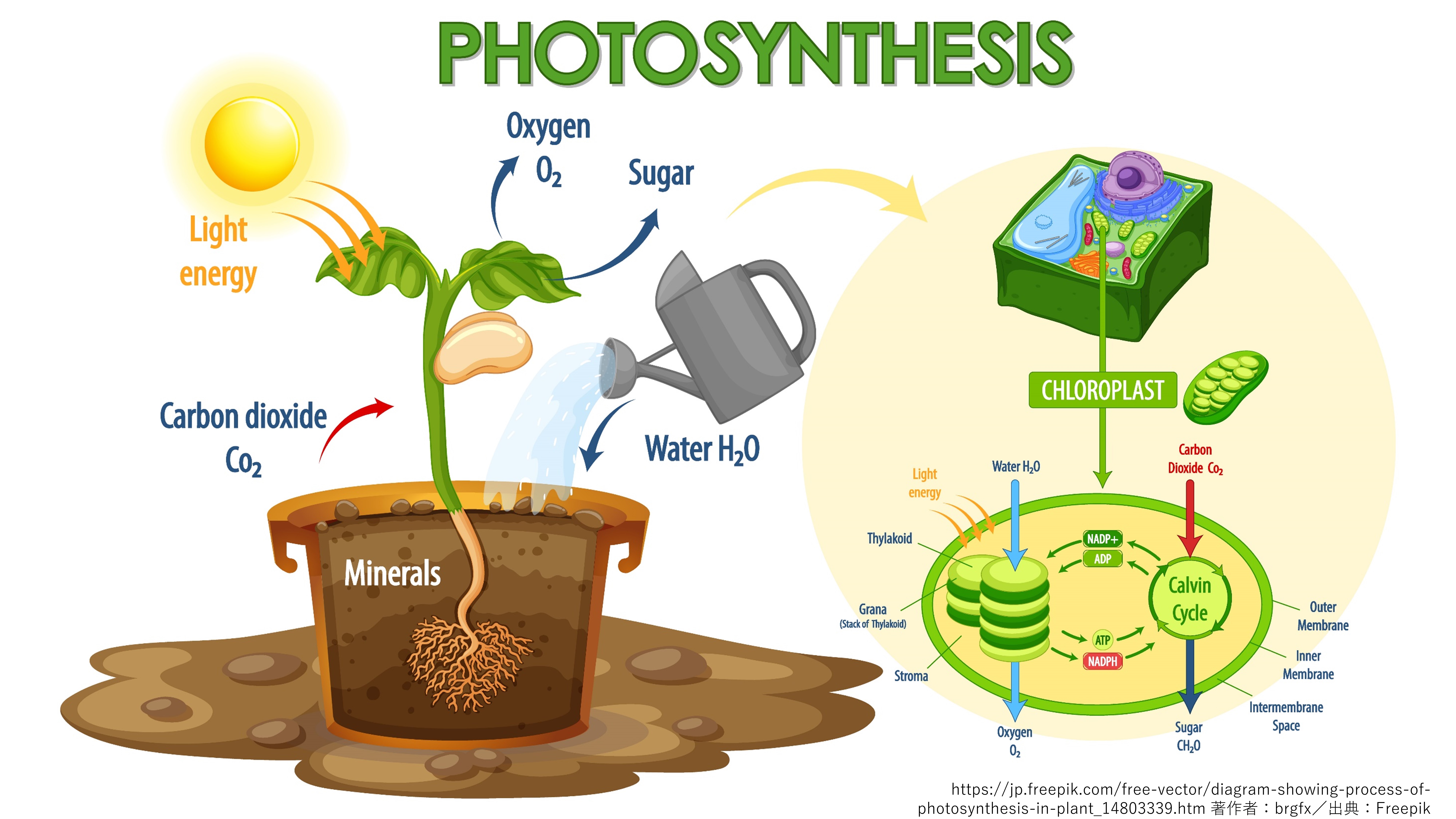
光合成は、太陽光等から得られるエネルギーを利用して無機炭素から有機化合物 (デンプン・糖等) を合成する反応を指します。光合成では、その過程で水が分解されて、副産物として酸素が放出されます。光合成は、地球上では陸上植物、藻類、及びシアノバクテリアによって行われます。
イネなどの作物は、光合成の活性度の大小によって生合成されるデンプン・糖の量が変化し、収穫高に差が出ます。つまり、作物の光合成能力を高めることにより、作物の増収が可能になります。
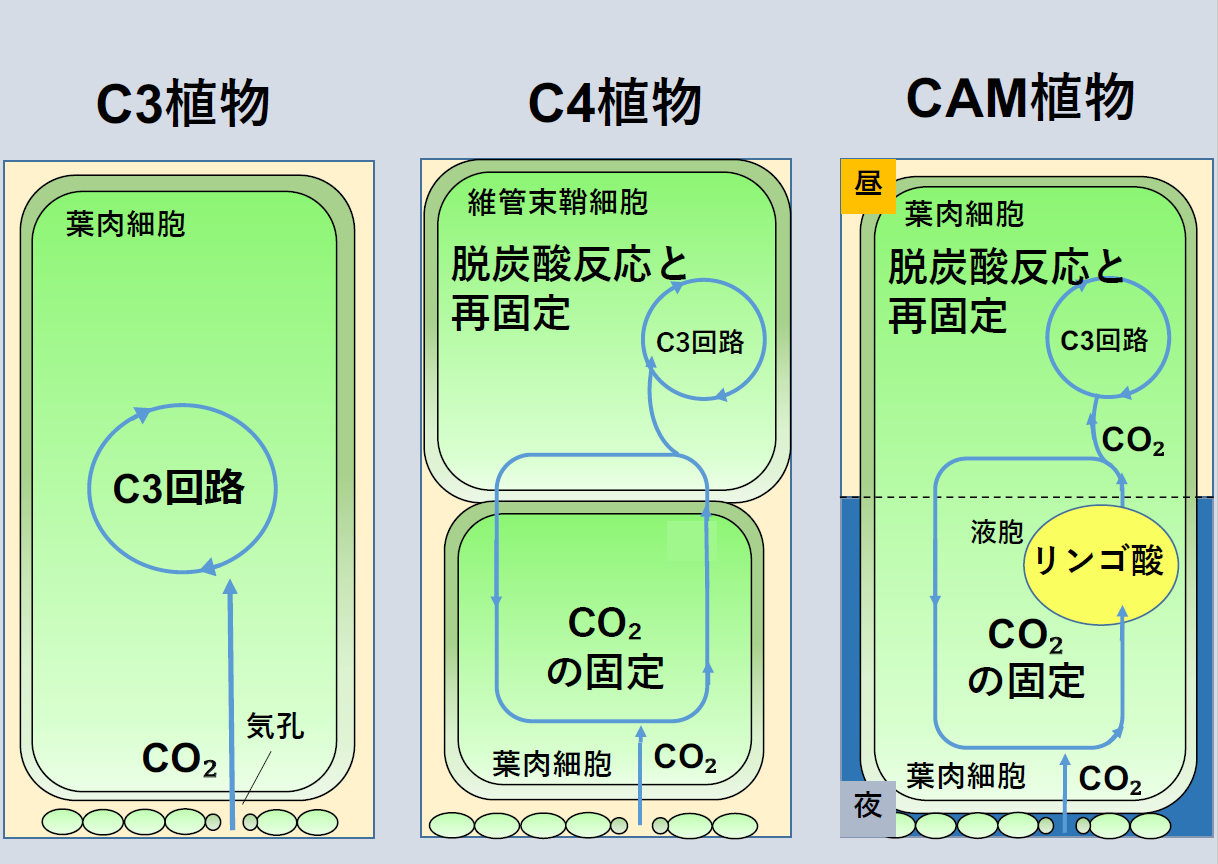
地球温暖化等の環境変動は、耕地環境を劣化させます。これによる、食糧生産性の低下は早急に解決すべき課題です。植物が有する光合成型は3種類に分けられます。イネ・ムギ等が有するC3型光合成、トウモロコシ・サトウキビ等が有するC4型光合成、並びにサボテン・アロエ等が有するベンケイソウ科有機酸代謝(Crassulacean
Acid Metabolism: CAM)型光合成に分類されます。主要作物の多くはC3型光合成を有する植物(C3植物)です。世界の耕地面積の約40%はこれらの生育に十分な降水量を満たしていないため、多くの食糧作物は常に環境ストレスに曝されている状態であると言えます。一方で、C4型及びCAM型光合成を有する植物(C4植物及びCAM植物)は乾燥・高温等の環境ストレスに対して高い耐性を示します。
そこで、C3型光合成を有するイネ等の作物に、C4型光合成またはCAM型光合成の形質を導入することで、環境ストレスに対してレジリエント(強靭)、すなわち高い耐性を有する作物の作出が可能になると期待されます。
また、地球上の植物の中には、耐塩性、耐乾性、及び耐冷性等の環境ストレス耐性を有するものが数多く存在します。私たちは、これらの植物の環境ストレス耐性機構にも着目し、生理学及び分子生物学的手法を駆使して、過酷な環境でも生きることができるメカニズムを明らかにする研究を進めています。
脱炭素社会の実現に向けた取組み
バイオマスエネルギーは、糖質、デンプン質、及び油脂等を豊富に含む作物を主体とするカーボンニュートラルな再生可能エネルギー資源です。その需要は近年増加しており、IEA(2021)によれば、2050年には全エネルギー需要の約2割を占めると推算されています。炭素排出量の低減による脱炭素社会を実現するために、エネルギー資源となる作物の開発が急務とされています。
私たちの研究室では、光合成型変換及び環境ストレス応答機構を導入した、エネルギーとしての側面を有する資源作物の開発も視野に入れています。バイオエタノールやバイオディーゼルを中心としたカーボンニュートラルエネルギーの研究は海外(特にアメリカ合衆国)では広く進められていますが、日本での研究例は少ないです。私たちは、原料となるデンプン等の光合成産物を効率よく素早く合成する作物の作出に向け研究に取り組んでいます。エネルギー分野の方々とのコラボレーションができれば嬉しいです。
取り扱う主な植物種
①アイスプラント (The common ice plant) (Mesembryanthemum crystallinumL.)

アイスプラントは、南アフリカ原産の一年生草本で、塩化ナトリウム(NaCl)存在下で生育が可能です。本種は、海水と同程度の濃度 (400 mM)
のNaCl存在下で生育すると、その光合成型をC3型からCAM型に変換します。また、他の作物が枯死する100 mM (海水の1/4程度) のNaCl環境下で生育を促進させます。塩存在下で生育する塩生植物の一種であり、他の塩生植物では種子に油脂を蓄積する報告がある(Yanxia,
2010)ことから、アイスプラントの種子にも油脂を蓄積する作用があることが示唆されています。
1970年代から本格的に本種の研究が始まり、今年度ゲノム配列が報告されました(Shen et al., 2022)。私たちの研究室でも独自にゲノム配列を構築し、ゲノムを用いたアイスプラントの分子生物学的解析が可能になっています。
②エレオカリス・ビビパラ (Umbrella hairgrass) (Eleocharis vivipara Link.)

エレオカリス・ビビパラはカヤツリグサ科ハイリ属の水陸両生植物で、生育環境の違いによりC3型光合成とC4型光合成を可逆的に変換します。本種には葉身がなく、稈が光合成器官として機能します。陸生型の稈は、葉緑体やミトコンドリアを多量に含んだ大型の維管束鞘細胞を有します。その外側には放射状に配列した柵状の葉肉細胞が取り囲み、C4植物に特有なクランツ型葉構造を示します。一方で、水中型の稈は表皮の内側に大型の発達した葉肉細胞を持ちます。維管束鞘細胞のサイズは著しく小さくなり、わずかな小型の葉緑体やミトコンドリアのみを含む細胞と化します。
また、エレオカリスでは陸生型と水生型で光呼吸の特性も異なっており、C3植物およびC4植物における光呼吸酵素の組織特異的な発現や光呼吸の制御機構及び生理的意義を研究する上でも有用な植物といえます。
③シュロガヤツリ (Umbrella plant) (Cyperus alternifolius L.)

シュロガヤツリは、湿地に生育するマダガスカル島原産のカヤツリグサ科カヤツリグサ属の多年生草本です。縣・宋(2002)は、本種によって富栄養化の進んだ水域の水質が改善されることを示し、環境浄化への有用性を明らかにしました。本種は、リンや窒素などの栄養素の吸収能力も高い(Kumwimba
et al., 2017)ことから、バイオマス資源としても有望です。さらに本種は、CrやPbなどの有害物質も吸収・蓄積します。
水生植物のバイオマスにはデンプン等の多糖類が多く含まれ、バイオエネルギーの原料になります。先行研究で、示差熱解析によって本種に含まれるバイオガスの量を推定したところ、ガス化効率が高く灰分が少ない事からバイオ燃料の原料となりうることが明らかにされました(田中,
2016)。
④イネ (Rice) (Oryza sativaL.)

イネはイネ科イネ属の植物で世界三大作物の一つです。近年アジアの人口増加により生産量が増えています。収量を上げるために化学肥料を使用することが多く、これは環境汚染や経済的な負担をもたらします。特に窒素は需要が多く、少ない施肥で高い収量を得ることが求められています
・C3植物へのCAM型光合成の付与
CAM植物は、細胞の構造がC3植物と類似していることが分かっており、C3植物の光合成代謝をCAM植物と同様のものに変換することが出来れば、C3植物にCAM型光合成の特性を付与することが可能になると示唆されています。私たちは、その仮説の実現性を実証し、環境ストレス耐性の高い作物を作出するために、遺伝子の発現量が転写因子によって制御される分子システムに着目した研究を行っています。
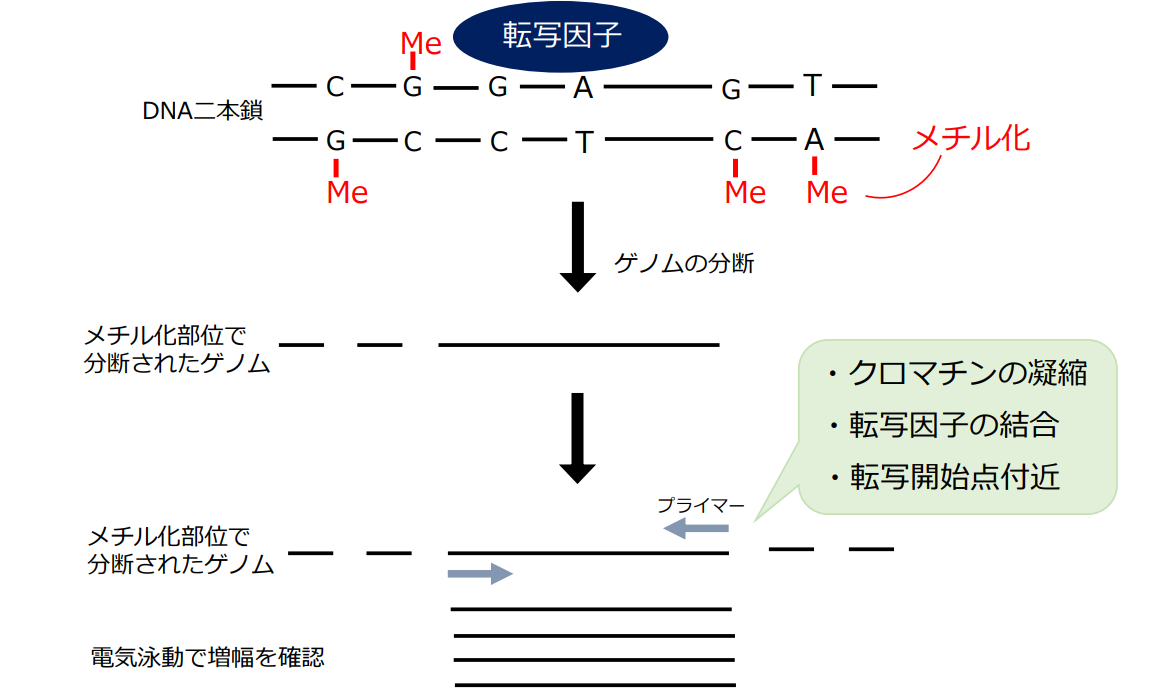
CAM型光合成は、朝にCO₂を吸収し、夜に有機物として固定する「時間的分業」を成功させています。先行研究では本研究で着目しているCAM型光合成の「時間的分業」に関わる遺伝子の発現量を継時的に制御する遺伝子の存在が示唆されました(Cushman et al., 2008)。この遺伝子が所謂「時計遺伝子」です。本研究では、CAM型光合成関連遺伝子の発現量をコントロールする転写因子を同定するために、ジメチル硫酸(DMS)を用いて転写因子結合部位を特定する独自の手法を用いた解析を進めています。DMSはタンパク質が結合した領域以外のDNAにメチル基を付与することで、分解を促します。そのため、結果的に転写因子が結合していたゲノム上の部位を推定することができます。
(担当)M2 近藤、宗岡、M1 Masako、高木、B4 岸本・環境要因の変動に伴うC3-C4光合成変換機構の解明

C4植物は、C3植物から長い年月をかけて進化してきたと言われています。植物の光合成型の進化は、約14億年前に始まった地球の大気CO2濃度低下が主要因と考えられています。しかしながら、その詳細な進化メカニズムは明らかにされておらず、多くの不明な点が残されています。ここでは、水陸両生植物エレオカリス(Eleocharis vivipara Link.)を用いて、C3型光合成からC4型光合成に進化するメカニズムを明らかにする研究を進めています。本種は、陸生型と水生型で異なる光合成型を示し、前者ではC4型、後者ではC3型光合成を行います。先行研究で、植物ホルモンが光合成型の変換に関与することが示唆されています。本研究では、栽培環境及び植物ホルモン量を変化させた処理区間で発現量が変動する遺伝子群をRNA-Seq解析によって網羅的に同定します。
(担当) D2 佐藤、M1 重政
・C3植物へのC4型光合成の導入法の開発
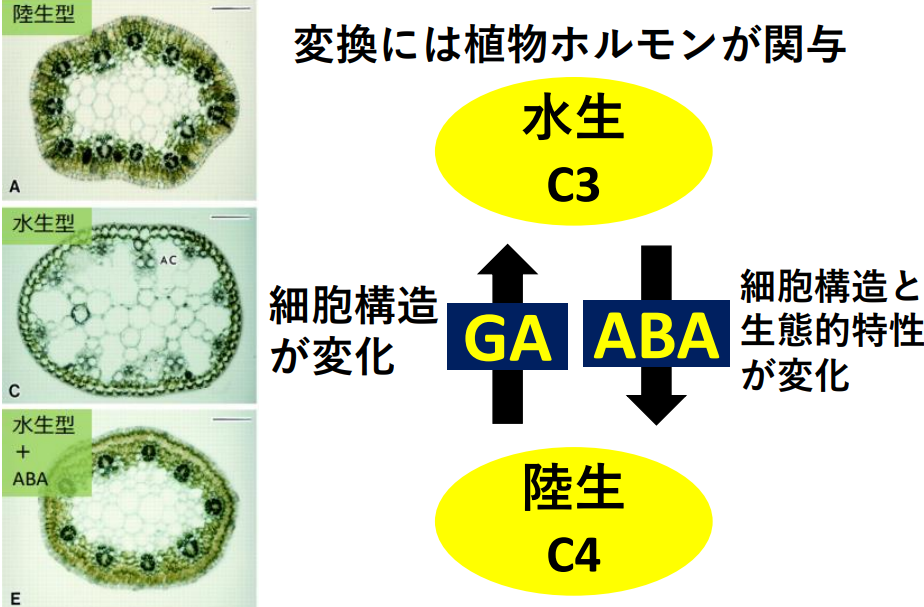
C3植物にC4光合成の代謝機構を導入する手法は、これまで多くの研究者によって試みられてきましたが、いずれも成功していません(Taniguchi et al., 2008; Miyao et al., 2011)。その原因の1つとしてC3植物及びC4植物の葉構造及び代謝機構が異なることが挙げられます。特に、C4植物がクランツ型葉構造を有することかつその形成過程が不明であることが、C3植物のC4化を困難にさせています。私たちの研究室では、このクランツ型葉構造を形成を促す遺伝子を解明する研究を進めており、その遺伝子を導入することでC3植物であるイネなどの主要作物にクランツ型葉構造を導入することができると考え、研究を進めています。
(担当) D2 佐藤、M1 重政
・低窒素条件下でのイネの高収量品種の作成

作物の収量を上げるために必要な化学肥料に含まれる窒素化合物は、河川に流出し水質汚染を引き起こすことが問題視されています。また、今後人口増加が予想される発展途上国などでは肥料は高価で十分に使えないために作物の増収が見込めないのが現状です。 収量を維持しつつこれらの問題を解決するには、肥料が少ない環境でも光合成能力を維持できる新品種の開発が必要です。そこで、低窒素条件下で高収量が得られるイネ品種の作出を目指し、窒素による光合成関連酵素遺伝子のエピジェネティックな発現制御機構について研究しています。また、形質転換技術の1つとしてゲノム編集を用いた実験も行います。
(担当) M2 宇都宮、M1 劉
・アイスプラントの地上部再分化の制御要因

アイスプラントには有効な形質転換体作出技術が確立されておらず、遺伝的な解析が困難です。この原因の一つに培養細胞から地上部が再分化しないことがあげられます。 この問題を解決するために、根外植体由来のカルスから地上部を再分化させる培養条件を検討しています。また再分化に関するに遺伝子の発現解析を行っています。これら二つの側面から、地上部が再分化しない原因を明らかにし、再分化法を確立させ、形質転換体の作出を試みています。
(担当) M1 神田
・アイスプラントの形質転換技術開発

アイスプラントの形質転換の効率的な方法は十分に確立されていません。そこで、私たちは多くの植物の形質転換に成功しているフローラルディップ法に着目しています。フローラルディップ法は、植物の蕾に直接Agrobacterium tumefaciensを感染させることで遺伝子変異を導入する手法です。
pBI7EGFPベクター等を保有するA. tumefaciens EHA105株を含む接種用落下培地を花に塗布し、GFPによる細胞局在性検出及び操作型顕微鏡による形態観察を介して、形質転換体を作出し選抜します。これまでの研究で、B培地(Luria Bertini base)よりフローラルディップ法に適したA培地(Murashige and Skoog base)を用いて、2つの健全な形質転換体を0.082
%で再現することに成功しています。また、花の発育段階は、スティグマの開き具合から、開花前3日目以降が最も適していることが示唆されています。
(担当) M2 Ginger
・アイスプラントの好塩性及びC3-CAM変換機構の解明

アイスプラントは海水の4分の1の濃度のNaCで処理すると生育が促進する「好塩性」を示す植物です。これまでにはキノアやスベリヒユなどで好塩性の研究が行われてきました。しかし、その分子生物学的メカニズムは不明なままです。
そこで、NaClがアイスプラントの生長にどのように関わっているかを、光合成やミトコンドリアのATP合成、細胞周期などに着目して研究を行っています。
また、本種は海水と同程度の濃度のNaCl存在下で自身の光合成型をC3型からCAM型に変換する機構を持ちます。このようなCAM植物を環境によって光合成型が変化しない偏性(Obligate)CAM植物に対して「通性(Fucultative)CAM植物」と呼びます。その生理学・形態学的現象は多くの研究者によって明らかにされてきましたが、偏性CAMと通性CAMの根本的な相違点であるCAM型光合成駆動機構は明らかにされていません。
本研究テーマでは、従来の作物学的・生理学的特性の調査に加え、バイオインフォマティクスを取り入れています。ゲノム、エピゲノム(DNAメチル化)、及びトランスクリプトームの観点から好塩性機構及びC3-CAM光合成変換機構を明らかにしようと試みています。
(担当) D2 佐藤、B4 森
・自然栽培を可能にする土壌微生物-植物相互作用の解明

持続可能な農業法の1つに「自然栽培」があります。これは、栽培時に化学肥料を一切使用せず、植物にとって有益な「土壌微生物」を有効利用しています。土壌微生物は、生ごみや廃材等をすみかとして増殖し、植物にとっての栄養をつくることが経験的にわかっていますが、その科学的根拠はいまだ不明です。
本研究では、一般的な農法(慣行農法)及び自然栽培法における土壌微生物に含まれる生物種をメタゲノム解析によって網羅的に解析します。最終的に、どのような微生物が自然栽培において有用であるかを明らかにします。
(担当) D1 笹原
・モリンガの普及に関する研究

モリンガ(Moringa oleifera)はワサビノキ科モリンガ属のインド原産の木本植物です。葉、塊根、種子、樹皮、莢、花、果実等が利用されています。これまでに、脂肪酸及びエステル類48種、炭化水素類56種、アルコール類37種、ケトン類32種、アルデヒド類その他69種等を含むことが示されています。心臓・循環器系への作用、腫瘍抑制、解熱、抗てんかん、抗炎症、抗胃潰瘍、抗痙攣、利尿、排尿、血圧降下、コレステロール低減、抗酸化、抗糖尿、肝機能保護、抗菌・抗真菌等が報告されています。2013年から有用成分を高め安定供給するための栽培法及び加工法の確立に取り組んでいます。2019年からは香川県三豊市で、2020年からは佐賀県唐津市で栽培試験を開始しました。共同研究募集中です。
(担当)M2 Joshua
・新機能性野菜としてのアイスプラントの利用

アイスプラント(Mesembryanthemum crystallinum L.)は南アフリカ原産の1年生草本です。本種はプロリンやピニトールといった機能性成分を持っています。この植物は様々な商標で全国で販売されていますが品種は野生のものと私たちが開発したKA-i243のみです。機能性を高めた品種の開発や機能性成分を高める栽培方法を確立することで新規機能性野菜としてアイスプラントを作成することを目的としています。
(担当)M1 Dennis
・シュロガヤツリを用いた水質浄化

シュロガヤツリ(Cyperus alternifolius L.)は、カヤツリグサ科カヤツリグサ属の植物です。下水処理の際に殺菌に用いられている次亜塩素酸ナトリウムをシュロガヤツリを用いて除去する手法を開発する研究を行っています。その一環で、シュロガヤツリが耐えられる次亜塩素酸ナトリウムの限界濃度を調査しています。また、養分吸収能力が著しく高く、富栄養化の改善にも役立つ植物であるため、富栄養化した水に浮かべた稈でシュロガヤツリを栽培し、水質調査及び生長量を調査しています。
また、シュロガヤツリの根系を観察すると、富栄養条件において繊毛が大量に増殖する現象が確認されています。本研究室では、共生細菌に焦点を当て、富栄養条件で生育が促進されるシュロガヤツリ及び周辺細菌の関連性を明らかにすることを目標としています。
(担当) D2 佐藤、M2 Joshua、M1 Dennis、B4 津留